


Tropical Forest
In collaboration with Corine Vriesendorp, The Field Museum of Natural History, Chicago. Check out the Field Museum guide to seedlings / virtual herbarium, including seedlings pictures taken by Corine in this project in Costa Rica.
This project has been funded by the National Science Foundation (NSF), Division of Environmental Biology, Long Term Research in Environmental Biology (LTREB).
Overview
 One of the major goals of the science of ecology is to understand the factors that determine the relative abundance and number of plant species in natural communities. Wet tropical forests contain a great diversity of tree species and provide a unique situation to test the factors that influence which tree species compose a forest. Survival and growth through the tree seedling stage is a critical bottleneck. The seedling phase of a plant’s life cycle is especially sensitive to pathogens and to the availability of light, nutrients, and water. Trees can live for much longer than humans and important events occur infrequently (e.g., some tree species produce seed at intervals of >12 years), necessitating longer-term studies. This research is based on a ~13 year record of seedling surveys in five forest stands in Costa Rica, with the goal of developing models of critical aspects of forest regeneration (seedling production, dispersion, growth and survival) in response to light, soil nutrients, pathogens, and rainfall.
One of the major goals of the science of ecology is to understand the factors that determine the relative abundance and number of plant species in natural communities. Wet tropical forests contain a great diversity of tree species and provide a unique situation to test the factors that influence which tree species compose a forest. Survival and growth through the tree seedling stage is a critical bottleneck. The seedling phase of a plant’s life cycle is especially sensitive to pathogens and to the availability of light, nutrients, and water. Trees can live for much longer than humans and important events occur infrequently (e.g., some tree species produce seed at intervals of >12 years), necessitating longer-term studies. This research is based on a ~13 year record of seedling surveys in five forest stands in Costa Rica, with the goal of developing models of critical aspects of forest regeneration (seedling production, dispersion, growth and survival) in response to light, soil nutrients, pathogens, and rainfall.
A better understanding of the factors that govern the relative abundance of tree species will aid in predicting forest responses to environmental changes, which is critical to timber production, wildlife habitat, and forest carbon storage. Environmental trends in tropical areas include greater variability in climate (warmer temperatures and less rainfall) and increasing deposition of nitrogen, sulfur, and other chemical pollutants from the atmosphere. This research will help resolve how forest regeneration responds to these environmental changes.
Recent Publications (Complete list of publications)
Minor, D and RK Kobe. 2019. Fruit production is influenced by tree size and size-asymmetric crowding in a wet tropical forest. Ecology and Evolution 9:1458-1472. https://doi.org/10.1002/ece3.4867
Holste, EK and RK Kobe. 2017. Tree species and soil nutrients drive tropical reforestation more than associations with mycorrhizal fungi. Plant and Soil 410: 283-297.
Holste, EK, RK Kobe, CA Gehring. 2017. Plant species differ in early seedling growth and tissue nutrient responses to arbuscular and ectomycorrhizal fungi. Mycorrhizae 27: 211-223.
Holste, EK, K Holl, Z Zahawi, RK Kobe. 2016. Reduced aboveground tree growth associated with higher arbuscular mycorrhizal fungal diversity in tropical forest restoration. Ecology and Evolution 6: 7253-7262.
Record, S, RK Kobe, CF Vriesendorp, and AO Finley. 2016. Seedling survival responses to conspecific density, soil nutrients, and irradiance vary with age in a tropical forest. Ecology 97: 2406-2415.
Bachelot, B, RK Kobe and CF Vriesendorp. 2015. Negative density-dependent mortality varies over time in a wet tropical forest advantaging rare species, common species, or no species. Oecologia 179: 853-861.
Kobe, RK, TB Baribault, and EK Holste 2014. Tree performance across gradients of soil resources. In Coomes DA, Burslem DFRP, and Simonson WD (eds.). Forests and Global Change, Ecological Reviews Series, Cambridge University Press, New York.
Guhaniyogi R., AO Finley, S Banerjee, and RK Kobe, 2013. Modeling complex spatial dependencies: low-rank spatially varying cross-covariances with application to soil nutrient data. Journal of Agricultural, Biological and Environmental Statistics 18: 274-298.
Bachelot, B and RK Kobe. 2013. Rare species advantage? Richness of damage types due to natural enemies increases with species abundance in a wet tropical forest. Journal of Ecology 101: 846-856.
Baribault, TB., RK Kobe, AO Finley. 2012. Tropical tree growth is correlated with soil phosphorus, potassium, and calcium, though not for legumes. Ecological Monographs 82: 189-203.
Holste, EK, RK Kobe, and CF Vriesendorp. 2011. Seedling growth responses to soil nutrients in a wet tropical forest understory. Ecology 92: 1828-1838.
Kobe, RK and CF Vriesendrop. 2011. Conspecific density-dependence in seedlings varies with species shade tolerance in a wet tropical forest. Ecology Letters 14: 503-510. doi: 10.1111/j.1461-0248.2011.01612.x
McCarthy-Neumann, S and RK Kobe. 2010. Conspecific plant-soil feedbacks reduce survivorship and growth of tropical tree seedlings. Journal of Ecology 98:396-407. doi: 10.1111/j.1365-2745.2009.01619.x
Kobe, RK and CF Vriesendorp. 2009. Size of sampling unit strongly influences detection of seedling limitation in a wet tropical forest. Ecology Letters 12: 220-228. doi: 10.1111/j.1461-0248.2008.01278.x
McCarthy-Neumann, S and RK Kobe. 2008. Tolerance of neighborhood soil pathogens co-varies with shade tolerance across species of tropical tree seedlings. Ecology 89:1883-1892.
de Gouvenain, RC, RK Kobe, and JA Silander, Jr. 2007. Partitioning of understory light and dry-season soil moisture gradients among seedlings of four rain-forest tree species in Madagascar, Journal of Tropical Ecology 23: 569-579.
Kobe, RK. 1999. Tropical tree richness and resource-based niches. Science.285: 1459a – 1460a.
Kobe, RK. 1999. Light gradient partitioning among tropical tree species through differential seedling mortality and growth. Ecology 80: 187-201.
Important Findings
While titles of the above publications usually summarize the important findings, a little more detail is provided here:
1. Spatial scale strongly influences estimates of fundamental seedling limitation (Kobe and Vriesendorp 2009). Fundamental seedling limitation is the lack of at least one seedling of a species in a defined area, which in turn limits mature tree species composition. In an area occupied by a mature tree, only one seedling needs to occur and then survive and grow to replace the canopy occupant to alleviate seedling limitation. Relatively common canopy tree species that occurred in only 15% of 1m2 sampling units had at least one seedling present in ~75% of 1x20 m sampling units (which approximates the crown diameter of a mature tree).

Figure 1. The sampling units in seedling limitation studies are often much smaller than the mature tree that a seedling might someday replace. (a) Transects used for seedling censuses consisted of 200 contiguous 1 m2 quadrats. The typical sampling unit used in studies of recruitment limitation is <=1 m2. However, the spatial scale of mature plants is much larger as illustrated for a 20-m crown diameter canopy tree. This study evaluated seedling prevalence at both 1 m2 and mature plant size spatial scales. (b) The spatial distribution of occupied 1-m2 quadrats will influence the calculation of prevalence at coarser spatial scales. In both transects, 5 of 40 1-m2 quadrats are occupied (prevalence1m = 0.125), but because occupied 1-m2 quadrats are clumped in the upper but broadly distributed in the lower transect, prevalence8m= 0.25 in the upper and 1.0 in the lower transect.
2. Irradiance and mineral nutrients have similar prevalence and magnitudes of effect on seedling height growth in the understory (Holste et al. 2011).
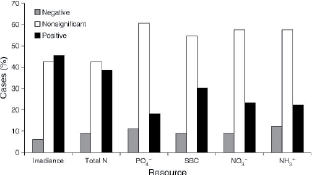
Figure 2. Percentage of species for which each resource was correlated with growth. SBC is the sum of base cations (potassium, magnesium, and calcium).
3. Species differences in (non-competitive) density-dependent mortality are more strongly related to physiologically based life history traits than biotic feedbacks originating from community abundance (Kobe and Vriesendorp 2011).
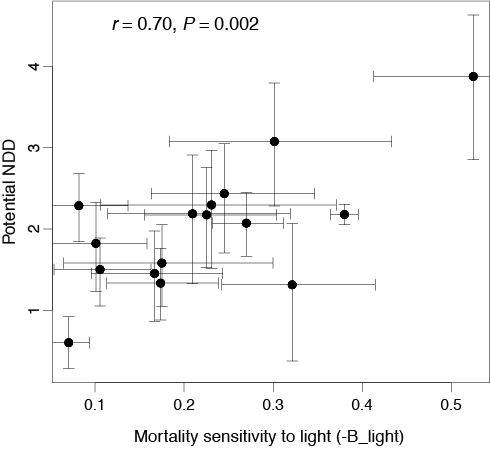
Figure 3. Across species, potential negative density dependence was strongly
correlated with sensitivity to shading (i.e. shade intolerance). Only species with
significant responses to irradiance and conspecific seedling density were included.
4. The diversity of damage types on leaves increases with local conspecific density (Bachelot and Kobe 2013).
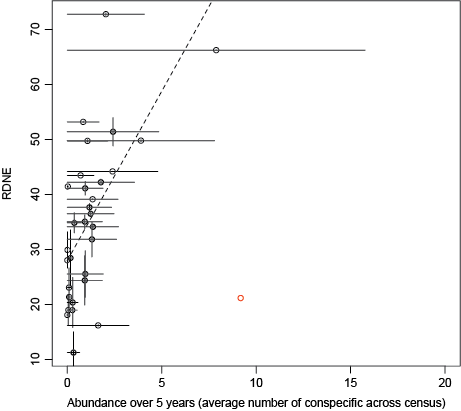
Figure 4. As the global abundance over the past 5 years increases, the
number of natural enemy species increases. Abundance is calculated
as the mean density of conspecific seedlings per m2. Vertical bars
represent the 95% confidence intervals. Pentaclethra macroloba is the
outlier in the lower right-hand corner. When P. macroloba is retained,
abundance over the past 5 year remains significant (P < 0.001,
R2 = 0.04 vs. P < 0.001, R2 = 0.3 when P. macroloba is excluded).
Site Description
 LaSelva is in the Atlantic lowlands of Costa Rica (10º 26’N, 84º 00’ W), with a mean annual rainfall of 4306 mm; during the January to April dry season, mean monthly minimum precipitation typically is > 100 mm. La Selva encompasses a broad gradient in soil fertility from relatively rich entisols and inceptisols of alluvial origin to extensive areas of low fertility ultisols developed on old lava flows (McDade and Hartshorn 1994). To represent landscape-level occurrence of major soils, we established three sites on residual soils and one site each on older and recent alluvial soils. Each of the five sites is centered on a belt transect of 200 contiguous 1m2 quadrats for monitoring seedlings. We mapped, measured diameters, and identified to species all trees > 5 cm diameter at breast height (at 1.37 m), originally in a 20 m buffer around the seedling belt transect. We added an additional 20 m buffer of mapped area to the core area around 2007-2009. Thus, each site is now 280 m length x 81 m width (Fig. 5).
LaSelva is in the Atlantic lowlands of Costa Rica (10º 26’N, 84º 00’ W), with a mean annual rainfall of 4306 mm; during the January to April dry season, mean monthly minimum precipitation typically is > 100 mm. La Selva encompasses a broad gradient in soil fertility from relatively rich entisols and inceptisols of alluvial origin to extensive areas of low fertility ultisols developed on old lava flows (McDade and Hartshorn 1994). To represent landscape-level occurrence of major soils, we established three sites on residual soils and one site each on older and recent alluvial soils. Each of the five sites is centered on a belt transect of 200 contiguous 1m2 quadrats for monitoring seedlings. We mapped, measured diameters, and identified to species all trees > 5 cm diameter at breast height (at 1.37 m), originally in a 20 m buffer around the seedling belt transect. We added an additional 20 m buffer of mapped area to the core area around 2007-2009. Thus, each site is now 280 m length x 81 m width (Fig. 5).

Fig. 5. Layout of each of the 5 sites at LaSelva Biological Station.
Core Data Set
Under support from the National Science Foundation (DEB 0075472, DEB 0743609) and Michigan State University, we have collected extensive data sets on seedling dynamics, resource availability, and mature tree dynamics. We are happy to share data from this project, unless plans for external use conflict with the PIs’ plans for publication. We also are happy to collaborate in data interpretation. Please contact Rich Kobe if you are interested in using data from this project. The major data sets are described below.
 Seedlings: we have monitored seedling establishment, height growth, and survival every six weeks from February 2000 through present in five 1 x 200m belt transects in wet tropical forest at La Selva Biological Station. As of August 2013, there are ~50,000 seedlings of ~400 species and morphospecies in our database. To our knowledge, this short census interval is unprecedented in a long-term study of early seedling establishment.
Seedlings: we have monitored seedling establishment, height growth, and survival every six weeks from February 2000 through present in five 1 x 200m belt transects in wet tropical forest at La Selva Biological Station. As of August 2013, there are ~50,000 seedlings of ~400 species and morphospecies in our database. To our knowledge, this short census interval is unprecedented in a long-term study of early seedling establishment.
In each seedling census at a given site, all newly emerging woody dicot (lianas, treelets, and subcanopy and canopy trees) and palm seedlings are marked with numbered tags and identified to species; previously tagged seedlings are monitored for survival. All individuals are measured for height. For each encountered species/ morphospecies, we take digital photographs as voucher specimens, which are archived in a virtual herbarium.
Resources: For each seedling quadrat, we have measured % canopy openness with hemispherical canopy photos in Fall 2000 and in July of 2004, 2006, and 2007 and in February and July of each year since 2008. For soil nutrient analysis, we took a composite of 3 subsamples of the upper 20 cm of mineral soil of each seedling quadrat and 60 additional points in the mapped stand in 2008, for a total of 260 soil samples per site. We analyzed for Mehlich III extractable phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg), and KCl-extractable inorganic N pools (NH4+ and NO3-).
Trees: We mapped, measured diameters, and identified to species all trees > 5 cm diameter at breast height (at 1.37 m), originally in a 20 m buffer around the seedling belt transect. Diameters have been measured and survival assessed on these trees in the following years: 2000 (although buttresses compromised some of the original measurements, 2005, and roughly every two years thereafter: 2007, 2009, 2011, and 2013.
Flowers and fruit production: We have censused all trees in the core area (41m x 240m) for flowering and fruiting four times: fall 2007, spring 2008, late 2009 – 2010, and 2012. We censused both the core area and the newly mapped zones in the two most recent censuses (2009-10 and 2012).
The numbers of flowers and /or fruits are estimated visually with binoculars and are placed into the following six categories: 0, 1-10, 11-100, 101, 1000, 1001 – 10,000, and > 10,000.
 Print
Print Email
Email